If you’re interested, please have a look at my book!
If you’re interested, please have a look at my book!
In previous articles I’ve written about the heath benefits of eating whole grains, which have been shown in large epidemiological studies to be associated with reduced all-cause mortality risk. Based on this data, the USDA’s MyPlate recommends a minimum of 3.5 oz, up to 7 oz. of whole grains on a 2200 calorie diet. 3.5 servings of barley yields 350 calories, whereas 7 oz. yields 700. In terms of percentage of total calories, MyPlate recommends that 16-32% of daily calories should come from whole grains.
In terms of vegetables, MyPlate’s recommendations are shown below. They recommend 3 servings of vegetables per day, with these amounts varied between green vegetables (and other vegetables), red and orange vegetables, beans and peas, and starchy vegetables. For ease of calculation I grouped ‘other vegetables’ with green vegetables. Based on the recommended weekly servings for each group and representative foods, I calculated weekly calorie amounts for each group. Average veggie calories per day = 187. Divided by 2200 calories, that equals 8.5% of total calories.

So clearly MyPlate wants us to eat between 2-4 fold more whole grains than veggies, in terms of total daily calories, but why is that? In a meta-analysis of 7 studies including 660,186 subjects, increased vegetable consumption is also associated with reduced mortality risk, as shown below:

Maybe whole grains are superior to veggies in terms of nutrient density? To see if that’s true, in the Table below I compared the nutrient composition of broccoli, spinach and romaine lettuce against barley (the king of grains for fiber), whole wheat spaghetti and oats. How do they compare in terms of macronutrients, when each has 100 calories? First, it should be obvious that to get 100 calories of veggies (see the serving column), you will eat significantly more food. To most, this will seem like a bad thing. But more chewing for the same amount of calories may end up in eating less, an important fact because of the worldwide explosion in obesity rates. Second, each of these veggies have 2-3 fold more protein and 3-4 fold more fiber than than whole grains. So far, veggies are far superior to whole grains.

What about vitamin content? As shown below, veggies crush whole grains for vitamin content. Whole grains are not better than veggies in terms of vitamin content for any category.

Maybe mineral content is better in whole grains? As shown below, they’re not. Veggies are much better in 9/10 mineral categories, with whole grains having marginally more selenium than veggies.

Based on these data, I have now dramatically increased my daily vegetable intake, while reducing my whole grain intake. Shown below is a snapshot of today’s veggie (and some other foods, too) intake, and it’s also important to mention that this amount is now representative of my daily vegetable intake. I haven’t eliminated whole grains, only minimized them.

My total veggie intake between carrots, beets, green peas, corn, asparagus and 1 pickle spear is 50.6 oz, or 1416 grams. Considering that 1 serving of vegetables = 80g, I ate 17.7 servings of veggies today. That amount is almost equal to what MyPlate recommends to eat in 1 week!
If you’re interested, please have a look at my book!
References:
Nutrition data from ndb.nal.usda.gov
Wang X, Ouyang Y, Liu J, Zhu M, Zhao G, Bao W, Hu FB. Fruit and vegetable consumption and mortality from all causes, cardiovascular disease, and cancer: systematic review and dose-response meta-analysis of prospective cohort studies. BMJ. 2014 Jul 29;349:g4490.

How much selenium is optimal for health? To address this question, I’ll examine the association between circulating levels of selenium with all-cause mortality risk. Then, I’ll identify a dietary selenium intake that corresponds to optimal plasma selenium levels. Let’s have a look!
A variety of studies have investigated associations between plasma (or serum) selenium with risk of death from all causes:
In a 9-year study of 1,389 older adults (average age, 65y) plasma selenium values less than 1.09 micromolar (uM) were associated with significantly increased all-cause mortality risk, when compared with values greater than 1.22 uM (Akbaraly et al 2005):

In a 6-year study of 1,042 older adults (average age, 76y), plasma selenium values less than 0.83 um were associated with significantly increased all-cause mortality risk, when compared with values greater than 1.0 uM (Lauretani et al. 2008):

In a 5-year study of 632 older women (average age, 74y), serum selenium values less than 1.38 uM were associated with significantly increased all-cause mortality risk, when compared with the other 3 quartiles (1.38 to 1.52, 1.53 to 1.67, and >1.68 uM; Ray et al. 2006):

In a 12-year study of 13,887 middle aged adults (average age, 45y), serum selenium values between 130-150 ng/mL (equivalent to 1.65-1.9 uM) were associated with reduced all-cause mortality risk (Bleys et al. 2006). Increased mortality risk was associated with serum selenium values less than 1.3 uM (~102 ng/mL):

These data were confirmed in the same cohort (16,008 adults) that were followed for an additional 2 years (14 years total; Goyal et al. 2013). Baseline serum selenium values greater than 1.4 uM were associated with significantly reduced all-cause mortality risk, compared with values less than1.38 uM.
Finally, in a 13-year study of 1,054 older adults (average age ~76y), elevated plasma levels of selenium (risk ratios were reported without the corresponding selenium concentration) were associated with significantly decreased all-cause mortality risk (Bates et al. 2011).
Studies that show weaker or no association between circulating values of selenium with all-cause mortality risk include Gonzalez et al. (2007) and Wei et al. (2004). In Gonzalez et al. (2007), serum selenium values greater than 1.26 uM were associated with decreased all-cause mortality risk for older women (average age, 76y), but not men, during a 4-year follow-up. However, Gonzalez et al. (2007) may have been underpowered to detect significant associations with mortality risk because of the small study size (215 total subjects). In Wei et al. (2004), a younger cohort (average age, 56y) of 1,115 subjects were followed for 15 years, and no association between serum selenium with all-cause mortality was found. However, only 4% of the population (~46 subjects) had serum selenium values greater than 1.19 uM, a finding that suggests that this study was additionally under-sized to detect significant associations.
Collectively, these studies suggest that circulating selenium values greater than at least 1.0 uM (and up to ~1.9 uM) are optimal for reducing all-cause mortality risk. What dietary intake of selenium can achieve these circulating values?
Shown below is the correlation between dietary selenium with serum selenium in 205 older adults (average age ~75y; González et al. 2006). Let’s start with the RDA selenium value for adults older than 19 years, 55 micrograms (mcg; Institute of Medicine, 2000). 55 mcg of dietary selenium is correlated with a serum selenium value of ~80 ug/L (~1 uM). In support of this correlation, a dietary selenium intake of ~47 ug/day has been shown to correlate with a circulating selenium concentration of 0.95 uM (Navarro et al. 1995). Based on the evidence already presented, eating only the RDA for selenium and achieving circulating selenium values less 1 uM would be associated with increased risk of death from all causes in 6 of the 7 studies! Based on its association with all-cause mortality risk, the RDA selenium value of 55 mcg/day is too low.

Determining which dietary selenium intake is optimal for maximally reduced all-cause mortality risk depends on how you interpret the literature. Four of previously mentioned studies showed circulating selenium values greater than 1.2 uM (95 ug/L) to be associated with reduced all-cause mortality risk. Based on the plot of González et al. (2006), ~130 ug of dietary selenium is necessary to achieve a circulating selenium concentration of 1.2 uM. Three studies showed decreased all-cause mortality risk at circulating selenium levels > 1.38 uM (110 ug/L). The dietary selenium intake that corresponds to that concentration is ~180 ug of selenium/day. Accordingly, a dietary selenium intake between 130-180 ug/day may be optimal for reducing all-cause mortality risk.
Which foods are selenium rich? Brazil nuts are the best dietary source of selenium, as 1 Brazil nut (4 g) contains 77 mcg of selenium (http://ndb.nal.usda.gov/ndb/foods/show/3641?fg=&man=&lfacet=&count=&max=&qlookup=&offset=&sort=&format=Full&reportfmt=other&rptfrm=&ndbno=&nutrient1=&nutrient2=&nutrient3=&subset=&totCount=&measureby=&_action_show=Apply+Changes&Qv=.04&Q6825=1&Q6826=1&Q6827=1). To achieve a dietary selenium intake between 130-180 mcg/day, every day I eat 1 or 2 Brazil nuts. It’s important to note that selenium toxicity can occur at intakes ~400 mcg (Food and Nutrition Board 2000), so keeping an eye on Brazil nut intake is probably a good idea.
If you’re interested, please have a look at my book!
References:
Akbaraly NT, Arnaud J, Hininger-Favier I, Gourlet V, Roussel AM, Berr C. Selenium and mortality in the elderly: results from the EVA study. Clin Chem. 2005 Nov;51(11):2117-23.
Bates CJ, Hamer M, Mishra GD. Redox-modulatory vitamins and minerals that prospectively predict mortality in older British people: the National Diet and Nutrition Survey of people aged 65 years and over. Br J Nutr. 2011 Jan;105(1):123-32.
Bleys J, Navas-Acien A, Guallar E.Serum selenium levels and all-cause, cancer, and cardiovascular mortality among US adults. Arch Intern Med. 2008 Feb 25;168(4):404-10.
Broome CS, McArdle F, Kyle JA, Andrews F, Lowe NM, Hart CA, Arthur JR, Jackson MJ. An increase in selenium intake improves immune function and poliovirus handling inadults with marginal selenium status. Am J Clin Nutr. 2004 Jul;80(1):154-62.
Food and Nutrition Board, Institute of Medicine. Selenium. Dietary reference intakes for vitamin C, vitamin E, selenium, and carotenoids. Washington, D.C.: National Academy Press; 2000:284-324.
González S, Huerta JM, Fernández S, Patterson EM, Lasheras C. Food intake and serum selenium concentration in elderly people.Ann Nutr Metab. 2006;50(2):126-31.
González S, Huerta JM, Fernández S, Patterson AM, Lasheras C. Homocysteine increases the risk of mortality in elderly individuals. Br J Nutr. 2007; 97:1138–1143.
Goyal A, Terry MB, Siegel AB. Serum antioxidant nutrients, vitamin A, and mortality in U.S. adults. Cancer Epidemiol Biomarkers Prev. 2013 Dec;22(12):2202-11.
Hurst R, Armah CN, Dainty JR, Hart DJ, Teucher B, Goldson AJ, Broadley MR, Motley AK, Fairweather-Tait SJ. Establishing optimal selenium status: results of a randomized, double-blind, placebo-controlled trial. Am J Clin Nutr. 2010 Apr;91(4):923-31.
Institute of Medicine. Food and Nutrition Board. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids. Washington, DC: National Academy Press, 2000.
Lauretani F, Semba RD, Bandinelli S, Ray AL, Ruggiero C, Cherubini A, Guralnik JM, Ferrucci L. Low plasma selenium concentrations and mortality among older community-dwelling adults: the InCHIANTI Study. Aging Clin Exp Res. 2008 Apr;20(2):153-8.
Navarro M, López H, Ruiz ML, González S, Pérez V, López MC. Determination of selenium in serum by hydride generation atomic absorption spectrometry for calculation of daily dietary intake. Sci Total Environ. 1995 Dec 15;175(3):245-52.
Ray AL, Semba RD, Walston J, Ferrucci L, Cappola AR, Ricks MO, Xue QL, Fried LP. Low serum selenium and total carotenoids predict mortality among older women living in the community: the women’s health and aging studies. J Nutr. 2006 Jan;136(1):172-6.
Swanson CA, Longnecker MP, Veillon C, Howe M, Levander OA, Taylor PR, McAdam PA, Brown CC, Stampfer MJ, Willett WC. Selenium intake, age, gender, and smoking in relation to indices of selenium status of adults residing in a seleniferous area. Am J Clin Nutr. 1990 Nov;52(5):858-62.
Wei WQ, Abnet CC, Qiao YL, Dawsey SM, Dong ZW, Sun XD, Fan JH, Gunter EW, Taylor PR, Mark SD. Prospective study of serum selenium concentrations and esophageal and gastric cardia cancer, heart disease, stroke, and total death. Am J Clin Nutr. 2004 Jan;79(1):80-5.
Is there a BMI that is associated with maximally reduced risk of death from all causes? Let’s have a look at the data!
In a meta-analysis of 19 studies that included 1,460,000 adults (median age, 58 years) a BMI between 20-25 kg/m2 was associated with maximally reduced all-cause mortality risk (Berrington de Gonzalez et al. 2010):

However, in a meta-analysis of 32 studies that included 197,140 older adults (65 years+), a BMI between 24 and 31 kg/m2 was associated with maximally reduced all-cause mortality risk (Winter et al. 2014). More specifically, a BMI between 26-26.9 kg/m2 was associated with maximally reduced all-cause mortality risk for never-smokers (Winter et al. 2014):

So what’s optimal for longevity in terms of BMI, is it 20-25 kg/m2, or potentially higher, as reported in older adults? For additional insight about the association between BMI with all-cause mortality, I looked up the published BMI data for centenarians:
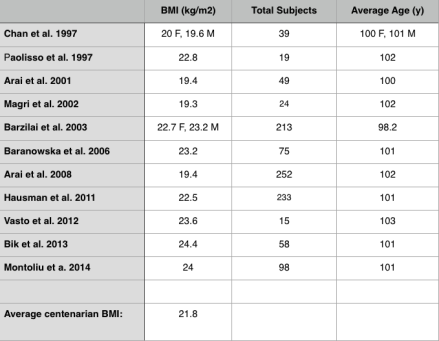
In these 11 studies that included 1075 centenarians, the BMI range was between 19.3-24.4 kg/m2, with an average BMI of 21.8. Shouldn’t that be the BMI reference range for those interested in living past 100?
If you’re interested, please have a look at my book!
References
Arai Y, Hirose N, Yamamura K, Shimizu K, Takayama M, Ebihara Y, Osono Y. Serum insulin-like growth factor-1 in centenarians: implications of IGF-1 as a rapid turnover protein. J Gerontol A Biol Sci Med Sci. 2001 Feb;56(2):M79-82.
Arai Y, Takayama M, Gondo Y, Inagaki H, Yamamura K, Nakazawa S, Kojima T, Ebihara Y, Shimizu K, Masui Y, Kitagawa K, Takebayashi T, Hirose N. Adipose endocrine function, insulin-like growth factor-1 axis, and exceptional survival beyond 100 years of age. J Gerontol A Biol Sci Med Sci. 2008 Nov;63(11):1209-18.
Baranowska B, Bik W, Baranowska-Bik A, Wolinska-Witort E, Szybinska A, Martynska L, Chmielowska M. Neuroendocrine control of metabolic homeostasis in Polish centenarians. J Physiol Pharmacol. 2006 Nov;57 Suppl 6:55-61.
Barzilai N, Atzmon G, Schechter C, Schaefer EJ, Cupples AL, Lipton R, Cheng S, Shuldiner AR. Unique lipoprotein phenotype and genotype associated with exceptional longevity. JAMA 2003;290:2030–40.
Berrington de Gonzalez A, Hartge P, Cerhan JR, Flint AJ, Hannan L, MacInnis RJ, Moore SC, Tobias GS, Anton-Culver H, Freeman LB, Beeson WL, Clipp SL, English DR, Folsom AR, Freedman DM, Giles G, Hakansson N, Henderson KD, Hoffman-Bolton J, Hoppin JA, Koenig KL, Lee IM, Linet MS, Park Y, Pocobelli G, Schatzkin A, Sesso HD, Weiderpass E, Willcox BJ, Wolk A, Zeleniuch-Jacquotte A, Willett WC, Thun MJ. Body-mass index and mortality among 1.46 million white adults. N Engl J Med. 2010 Dec 2;363(23):2211-9. doi: 10.1056/NEJMoa1000367. Erratum in: N Engl J Med. 2011 Sep 1;365(9):869.
Bik W, Baranowska-Bik A, Wolinska-Witort E, Kalisz M, Broczek K, Mossakowska M, Baranowska B. Assessment of adiponectin and its isoforms in Polish centenarians. Exp Gerontol. 2013 Apr;48(4):401-7.
Chan YC, Suzuki M, Yamamoto S. Dietary, anthropometric, hematological and biochemical assessment of the nutritional status of centenarians and elderly people in Okinawa, Japan. J Am Coll Nutr. 1997 Jun;16(3):229-35.
Hausman DB, Johnson MA, Davey A, Poon LW. Body mass index is associated with dietary patterns and health conditions in georgia centenarians. J Aging Res. 2011;2011:138015.
Magri F, Muzzoni B, Cravello L, Fioravanti M, Busconi L, Camozzi D, Vignati G, Ferrari E. Thyroid function in physiological aging and in centenarians: possible relationships with some nutritional markers. Metabolism. 2002 Jan;51(1):105-9.
Montoliu I, Scherer M, Beguelin F, DaSilva L, Mari D, Salvioli S, Martin FP, Capri M, Bucci L, Ostan R, Garagnani P, Monti D, Biagi E, Brigidi P, Kussmann M, Rezzi S, Franceschi C, Collino S. Serum profiling of healthy aging identifies phospho- and sphingolipid species as markers of human longevity. Aging (Albany NY). 2014 Jan;6(1):9-25.
Paolisso G, Ammendola S, Del Buono A, Gambardella A, Riondino M, Tagliamonte MR, Rizzo MR, Carella C, Varricchio M. Serum levels of insulin-like growth factor-I (IGF-I) and IGF-binding protein-3 in healthy centenarians: relationship with plasma leptin and lipid concentrations, insulin action, and cognitive function. J Clin Endocrinol Metab. 1997 Jul;82(7):2204-9.
Vasto S, Scapagnini G, Rizzo C, Monastero R, Marchese A, Caruso C. Mediterranean diet and longevity in Sicily: survey in a Sicani Mountains population. Rejuvenation Res. 2012 Apr;15(2):184-8.
Winter JE, MacInnis RJ, Wattanapenpaiboon N, Nowson CA. BMI and all-cause mortality in older adults: a meta-analysis. Am J Clin Nutr. 2014 Apr;99(4):875-90.
Vitamin K is found in 2 predominant forms, Vitamin K1 (phylloquinone), found almost exclusively in green leafy vegetables, and Vitamin K2 (Menaquinone), found in fermented foods, organ meats, meat, butter and eggs. In the data below (Juanola-Falgarona et al. 2014), we see that Vitamin K1 (phylloquinone) is negatively associated with death from all causes:
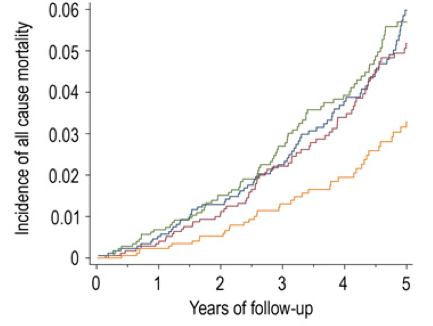
Death from all causes was assessed based on the average value for four groups of Vitamin K1 intake: 171 ug/day = blue line, 276 ug/day =red line, 349 ug/day = green line and 626 ug/day = the yellow line. In the data above, Vitamin K1 values less than 349 ug/day are about the same in terms of all-cause mortality risk. However, those who ate 626 ug/day of Vitamin K1 had about half of the mortality risk compared to the lower K1 intake groups! Interestingly, the RDA for Vitamin K, at 90 ug/day seems to be outdated, based on the data above. Also, Vitamin K2 was not associated with all-cause mortality risk, as shown below:

Based on the K1 mortality data, 626 ug/day seems like a good goal. However, osteocalcin is a Vitamin K-dependent protein that has been shown to be maximally active in the presence of 1000 ug of Vitamin K1 (Binkley et al. 2002)! Osteocalcin is involved in pathways that decline with aging: insulin secretion and β-cell proliferation in the pancreas, energy expenditure by muscle, insulin sensitivity in adipose tissue, muscle and liver, and increased testosterone production (Karsenty and Ferron 2012). Therefore, getting 1000 ug+ per day of Vitamin K1 may optimize all of these functions and, decrease mortality risk!
What’s the take home from these data? Eat more leafy greens! How much is needed to get 1000 ug per day? Shown below is a short list of foods rich in Vitamin K and the serving size needed to reach 1000 ug. Approximately 4 ounces of cooked kale or 7 oz. of raw spinach will suffice, and at a low calorie yield. Other foods, like broccoli, brussel sprouts or romaine lettuce would need to be consumed in far greater amounts to reach 1000 ug.

What’s my daily K1 intake? Shown below is my 7-day average (7/16/2015 – 7/22/2015) for K intake, derived almost exclusively from plant sources. 1379 ug/day puts me well above the 626 ug/day that was associated with reduced mortality risk, and above the 1000 ug/day needed for maximal osteocalcin activation.

If you’re interested, please have a look at my book!
References:
Binkley NC, Krueger DC, Kawahara TN, Engelke JA, Chappell RJ, Suttie JW. A high phylloquinone intake is required to achieve maximal osteocalcin gamma-carboxylation. Am J Clin Nutr. 2002 Nov;76(5):1055-60.
Juanola-Falgarona M, Salas-Salvadó J, Martínez-González MÁ, Corella D, Ostrich R, Ros E, Fitó M, Arós F, Gómez-Gracia E, Fiol M, Lapetra J, Basora J, Lamuela-Raventós RM, Serra-Majem L, Pintó X, Muñoz MÁ, Ruiz-Gutiérrez V, Fernández-Ballart J, Bulló M. Dietary intake of vitamin K is inversely associated with mortality risk. J Nutr. 2014 May;144(5):743-50.
Karsenty G, Ferron M. The contribution of bone to whole-organism physiology. Nature. 2012 Jan 18;481(7381):314-20.
Calorie restriction is well documented as the gold standard in terms of minimizing disease risk and maximizing longevity in almost every organism tested-worms, flies, mice, rats, dogs and monkeys. However, eating 10-30% less calories than usual is a difficult task for most people, as evidenced by the continuous rise in global obesity. But, there may be a way to reap the benefits of a calorie restricted diet without actually eating less calories, one that involves eating less methionine.
Methionine is one of the 20 essential amino acids, meaning that the human body cannot synthesize it, and therefore, it must be supplied by the diet. As shown below, a 5.5-fold restriction in dietary methionine (0.17% of total calories compared with 0.86%) without any other dietary changes (in rats) has been shown to increase average lifespan by ~20%, and maximal lifespan by 12% (Orentreich et al. 1993). Translated into human lifespan, a 20% increase would equate to an average lifespan from 75 to 90 years and a maximal lifespan from 122 to 134 years!

More importantly, food intake when normalized to body weight was greater in methionine-restricted animals, evidence that indicates that the lifespan extending effect was not due to a reduction in calories.

In other words, the methionine-restricted rats ate more than rats on the normal diet, but, these calories were not deposited as fat (or muscle), but burned as heat. This concept, of an increase in heat production (as opposed to energy production) is known as uncoupling, and has also been shown to be associated with an increased lifespan (Speakman et al. 2004). Furthermore, dietary methionine restriction has been shown to increase uncoupling (Hasek et al. 2010), and may be playing a part in the observed extended lifespan shown by Orentreich et al. (1993).
So how can we incorporate methionine restriction into our every day diet? The easy answer would be to reduce overall protein intake. For example, a diet that included a 4-egg white omelet for breakfast, a tuna sandwich for lunch, and a relatively lean (85-15, protein-fat) burger for dinner would contain a total of ~75 grams of protein and ~2.1 grams of methionine (based on the methionine content list as reported by McCarty et al. 2009) . Replacing these protein (and methionine) rich sources with an equivalent amount of calories (80 calories, egg whites; 170 calories, tuna; 250 calories beef) with whole wheat pasta (500 calories, or any other grain) reduces the overall protein intake to 24 grams, with ~300 mg (0.3 grams) of methionine. As you can see, elimination of egg/fish/meat reduces the methionine content by 7-fold, and, may be feasible in humans as a means for increasing lifespan.
My 7-day average protein intake is shown below. Within that, my average methionine intake is 0.8g/day. However, it is important to note that the nutrient-tracking software that I use (cronometer.com) for some reason doesn’t have the amino acid breakdown for my daily can of sardines, which adds 0.6 g of methionine. In total, I consume on average, 0.8 g + 0.6 g = 1.4 daily grams of methionine. Each gram of protein contains 4 calories. Therefore, my daily methionine intake =1.4*4 = 5.6 calories. My average calorie intake during that 7-day period was 2241 calories. 5.6/2241 *100 = 0.25%, which puts me closer to the 0.17% long-lived diet than to the shorter-lived 0.86% of Orentreich et al. 1993.

If you’re interested, please have a look at my book!
References:
Hasek BE, Stewart LK, Henagan TM, Boudreau A, Lenard NR, Black C, Shin J, Huypens P, Malloy VL, Plaisance EP, Krajcik RA, Orentreich N, Gettys TW. Dietary methionine restriction enhances metabolic flexibility and increases uncoupled respiration in both fed and fasted states. Am J Physiol Regul Integr Comp Physiol. 2010 Sep;299(3):R728-39.
McCarty MF, Barroso-Aranda J, Contreras F. The low-methionine content of vegan diets may make methionine restriction feasible as a life extension strategy. Med Hypotheses. 2009 Feb;72(2):125-8.
Orentreich N, Matias JR, DeFelice A, Zimmerman JA. Low methionine ingestion by rats extends life span. J Nutr. 1993 Feb;123(2):269-74.
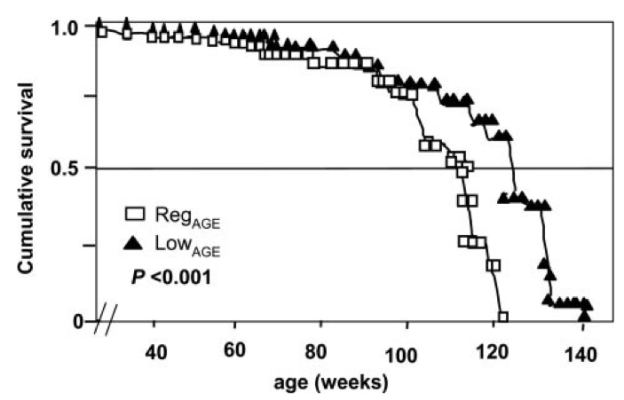
Dietary advanced glycation end products (AGEs) are formed as a result of the heat-induced (greater than 100C) binding of sugar to protein, fat or nucleic acids (DNA or RNA). The common way to identify AGE products in food is the browning effect: deep-frying, broiling, roasting, and grilling each produce a temperature that is sufficient to greatly increase AGE product formation, relative to either raw or boiled food.
The importance of dietary AGE products is that they shorten lifespan! Cai et al. (2007) quantified the amount of one particular type of AGE product, CML (carboxy-methyl-lysine) found in the mouse diet, and then fed mice half of this amount. The low-CML diet was sufficient to significantly extend median and maximal lifespan by 15% and 6%, respectively:
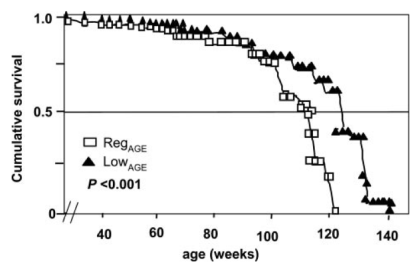
As shown below (inset), no difference in food intake was observed when comparing the 2 groups-from this it can be concluded that the lifespan extending effect of the low-CML diet was not related to a reduction in calorie intake. Also, mice on the low-CML diet had significantly decreased body weight, evidence that shows that it isn’t just calories that we should be worried about in terms of body weight maintenance.

Dietary AGE products also shorten lifespan on a calorie-restricted diet. Calorie restriction is the gold standard in terms of minimizing disease risk and extending longevity in a variety of organisms, including worms, flies, mice, dogs, and monkeys. Because CR mice eat less food than controls, the possibility existed that CR-fed animals also ate less AGE products. To address this possibility, Cai et al. (2008) quantified the amount of AGE products that CR-fed mice consumed, and then increased this amount to either equal to or greater than what mice on a regular diet ate. In terms of lifespan, mice on a low AGE, low calorie diet had increased average and maximal lifespan, relative to mice on a regular, ad libitum diet. However, lifespan was significantly reduced for CR-fed mice whose food was supplemented an AGE product amount that was equal to the regular diet!

These data suggest that if you eat less calories than normal, you will live longer, but, if your lower-calorie diet is poor in quality (i.e. high in AGE products), you will lose the lifespan extending effect of CR.
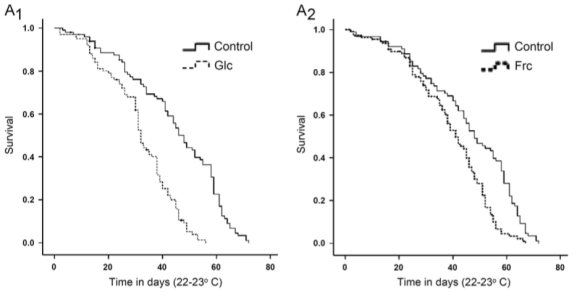
In support of the hypothesis that AGE products are bad for lifespan, dietary supplementation with glycated albumin (left) and fructosylated albumin (right) also shorten lifespan, in flies (Tsakiri et al. 2013):
To illustrate how cooking food at a high temperature impacts AGE formation, shown below is the AGE product (CML) content for a variety of foods (Goldberg et al. 2004):

For example, boiling beef (as in a chili recipe), compared with roasting it results in ~3-fold less AGE products. Boiling egg yolks results in about half as much CML when compared with frying. Interestingly, olive oil has more than double the amount of CML, when compared to broiled chicken or beef! Finally, fruits and vegetables such as bananas, apples, carrots, and green beans have almost negligible amounts of CML.
This post is now in video form!
References:
Cai W, He JC, Zhu L, Chen X, Wallenstein S, Striker GE, Vlassara H. Reduced oxidant stress and extended lifespan in mice exposed to a low glycotoxin diet: association with increased AGER1 expression.Am J Pathol. 2007 Jun;170(6):1893-902.
Cai W, He JC, Zhu L, Chen X, Zheng F, Striker GE, Vlassara H. Oral glycotoxins determine the effects of calorie restriction on oxidant stress, age-related diseases, and lifespan. Am J Pathol. 2008 Aug;173(2):327-36.
Goldberg T, Cai W, Peppa M, Dardaine V, Baliga BS, Uribarri J, Vlassara H. Advanced glycoxidation end products in commonly consumed foods. J Am Diet Assoc. 2004 Aug;104(8):1287-91.
Tsakiri EN, Iliaki KK, Höhn A, Grimm S, Papassideri IS, Grune T, Trougakos IP. Diet-derived advanced glycation end products or lipofuscin disrupts proteostasis and reduces life span in Drosophila melanogaster. Free Radic Biol Med. 2013 Dec;65:1155-63.
If you’re interested, please have a look at my book!
Our story begins with Michael Rose, who used an experimental evolution approach to breed flies that live more than 2-fold longer than controls (Rose 1999). In contrast with the one gene at a time knockout or overexpression strategy that is ubiquitous in modern biology, Rose separated initially genetically homogeneous flies into two groups-one with delayed and the other with normal reproduction. As shown below, after 80 generations, the group with continually delayed reproduction had ~2-fold increased average and maximal lifespan.

While this suggests that continually delaying reproduction may extend lifespan in people, the time it would take to do that makes it an unreasonable strategy. In 2010, the average age at first reproduction in the US was 25.4 years (http://www.cdc.gov/nchs/data/nvsr/nvsr61/nvsr61_01.pdf). Therefore, to replicate the doubling of lifespan found in Drosophila, 80 generations * 25.4y would take more than 2000 years! In contrast, a more reasonable approach towards extending life in people may involve stimulation of some of the pathways involved in the extended fly lifespan.
What genetic mechanisms underlie this 2-fold increase in Drosophila lifespan? Kurapati et al. 2000) found that levels of the mitochondrially located heat shock protein 22 (Hsp22) were between two and ten-fold higher in long-lived Drospohila, relative to the shorter-lived controls. In 2004 Morrow et al. identified the causative role of Hsp22 overexpression, as average lifespan increased by ~30%, thereby implicating mechanisms related to upregulation of Hsp22 on increasing lifespan.
Unfortunately, the role of Hsp22 on influencing lifespan in mammals is unknown. But, can we learn something about the underlying mechanism of the Hsp22-induced increase in lifespan and apply that to mammalian aging?
Hsp22 expression is regulated by histone deacetylase (HDAC) inhibitors (Zhou et al. 2005). In other words, when certain HDAC’s are inhibited, histone acetylation increases, resulting in elevated Hsp22 expression. Interestingly, histone acetylation has also been shown to be involved in lifespan determination in yeast and worms (Kaeberlein et al. 1999, Kang et al. 2002, Kim et al. 1999, Tissenbaum and Guarente, 2001). In contrast, the HDAC’s that have been popularized by the resveratrol-sirtuin story results in histone deacetylation, or the removal of acetyl groups from histones.
Identification of compounds that inhibit the HDAC’s that control Hsp22 expression would seem to be a good method for potentially increasing mammalian lifespan. Supplementation with sodium butyrate increases Hsp22 expression in Drosophila (Zhao et al. 2005), resulting in increased Drosophila lifespan (McDonald et al. 2013). Interestingly, sodium butyrate is a class I, II, IV HDAC inhibitor, whereas the sirtuins are class III inhibitors (Witt et al. 2009), evidence that suggests differing roles for the HDACs on lifespan extension.
Unfortunately, the causative role of butyrate-producing bacteria on mammalian lifespan has yet to be directly tested. However, butyrate stimulates expression of fibroblast growth factor 21 (Li et al. 2012), a protein whose overexpression extends both average and maximal lifespan in mice (shown below; Zhang et al. 2012).

Furthermore, acarbose supplementation extends median and maximal lifespan in genetically heterogeneous mice (Harrison et al. 2013), an important finding because acarbose supplementation has been shown to elevate serum butyrate in human subjects (Wolever and Chiasson 2000).
How can we get butyrate into our diet? Although butter contains small amounts of butyrate, a butter-rich diet has been shown to be obesogenic (Hariri et al. 2010). Fortunately, there is another way we can increase levels of butyrate, and that’s by stimulating our intestinal bacteria to produce it! The most abundant butyrate-producing gut bacterial species are Faecalibacterium prausnitzii, Eubacterium rectale, Eubacterium hallii and Anaerostipes hadrus (Tap et al. 2009, Walker et al. 2011).
Interestingly, butyrate-producing bacteria decrease during aging, which, in my opinion makes colonizing our intestines with these beneficial bacteria all the more important. For example, Faecalibacterium prausnitzii, Eubacterium hallii and Eubacterium rectale are significantly reduced in centenarians when compared with elderly and young subjects (Biagi et al. 2010). Several other gut bacterial butyrate producers are also reduced in centenarians when compared with the other age groups, including Ruminococcus obeum, Roseburia intestinalis, E. ventriosum, and, Papillibacter cinnamovorans.
Collectively, these data suggest that increasing gut bacterial species that produce butyrate may be important for increasing lifespan in both lower organisms and, in mammals. How can we boost butyrate-producing bacteria? Prebiotics, food ingredients that stimulate the growth and/or activity of bacteria in the digestive system may be the best option. Two such food components are inulin and fructooligosaccharides (FOS), which in vitro, stimulate growth of the butyrate producers F. prausnitzii, E.rectale, E. hallii and R. intestinalis 4-15 fold above basal levels (Scott et al. 2014). In vivo, consumption of 10g/day of inulin for 16 days in healthy, middle aged humans (BMI 25 kg*m-2, avg. age 38) significantly stimulated growth of F. prausnitzii (Ramirez-Farias et al. 2009). Therefore, consumption of foods rich in inulin and FOS may be a valid strategy for boosting levels of butyrate-producing bacteria in our intestines.
Foods containing inulin and fructoligosaccharides are shown in Table 1.

First, it’s important to mention that average intake for inulin and FOS is only ~2.5 grams/day (5g total) (Moshfegh et al. 1999). This is because relatively few foods contain high amounts of these prebiotic fibers. For example, whereas 100g of bananas (equivalent to 1 small banana) contains 1 gram total of combined inulin and FOS, in contrast, chicory root and Jerusalem artichoke contain 64.5 and 31.5 grams, respectively. In addition, although not shown in the table, most fruits contain limited amounts of FOS. Bananas contain more FOS/g than apple, blackberry, blueberry, cantaloupe, grapes, orange, peach, pear, raspberry, rhubarb, strawberry and watermelon (Dumitiriu 2005). This is an important finding because one would expect fruits to be rich in inulin and FOS, as both of these fibers contain long chain fructose polymers. Furthermore, based on values for asparagus, chicory, onion, loss of inulin and FOS upon boiling is ~30%, so eating these foods raw is not the only strategy for increasing dietary amounts of FOS and inulin.
Can increasing consumption of FOS and inulin improve health and lifespan? To date, dietary supplementation with inulin has been shown to improve cognitive performance (Messaoudi et al. 2005), and, to reduce cholesterol, triglycerides and body weight, and, improved survival in rats (Rozan et al. 2008). Although randomized controlled trials examining the effect of increasing butyrate-producing bacteria on health and mortality risk in older adults has yet to be performed, collectively, the evidence presented here suggests that if you’re interested in a low risk, potentially high reward approach towards improving health and lifespan, consuming more foods containing FOS and inulin may be a valid strategy!
If you’re interested, please have a look at my book!
References
Biagi E, Nylund L, Candela M, Ostan R, Bucci L, Pini E, Nikkïla J, Monti D, Satokari R, Franceschi C, Brigidi P, De Vos W. Through ageing, and beyond: gut microbiota and inflammatory status in seniors and centenarians. PLoS One. 2010 May 17;5(5):e10667.
Dumitiriu S. Polysaccharides: Structural Diversity and Functional Versatility. 2005. CRC Press. p854-855.
Hariri N, Gougeon R, Thibault L. A highly saturated fat-rich diet is more obesogenic than diets with lower saturated fat content. Nutr Res. 2010 Sep;30(9):632-43.
Harrison DE, Strong R, Allison DB, Ames BN, Astle CM, Atamna H, Fernandez E, Flurkey K, Javors MA, Nadon NL, Nelson JF, Pletcher S, Simpkins JW, Smith D, Wilkinson JE, Miller RA. Acarbose, 17-α-estradiol, and nordihydroguaiaretic acid extend mouse lifespan preferentially in males. Aging Cell. 2013 Oct 26.
Kaeberlein, M., McVey, M. and Guarente, L. (1999). The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 13, 2570-2580.
Kang, H. L., Benzer, S. and Min, K. T. (2002). Life extension in Drosophila by feeding a drug. Genetics 99, 838-843. Kim, S., Benguria, A., Lai, C. and Jazwinski, S. M. (1999). Modulation of lifespan by histone deacetylase genes in Saccharomyces cerevisiae. Mol. Biol. Cell 10, 3125-3156.
Kurapati R, Passananti HB, Rose MR, Tower J. Increased hsp22 RNA levels in Drosophila lines genetically selected for increased longevity. J Gerontol A Biol Sci Med Sci. 2000 Nov;55(11):B552-9.
Li H, Gao Z, Zhang J, Ye X, Xu A, Ye J, Jia W. Sodium butyrate stimulates expression of fibroblast growth factor 21 in liver by inhibition of histone deacetylase 3. Diabetes. 2012 Apr;61(4):797-806.
McDonald P, Maizi BM, Arking R. Chemical regulation of mid- and late-life longevities in Drosophila. Exp Gerontol. 2013 Feb;48(2):240-9.
Messaoudi M, Rozan P, Nejdi A, Hidalgo S, Desor D. Behavioural and cognitive effects of oligofructose-enriched inulin in rats. Br J Nutr. 2005 Apr;93 Suppl 1:S27-30.
Moshfegh AJ, Friday JE, Goldman JP, Ahuja JK. Presence of inulin and oligofructose in the diets of Americans. J Nutr. 1999 Jul;129(7 Suppl):1407S-11S.
Morrow G, Samson M, Michaud S, Tanguay RM. Overexpression of the small mitochondrial Hsp22 extends Drosophila life span and increases resistance to oxidative stress. FASEB J. 2004 Mar;18(3):598-9.
Ramirez-Farias C, Slezak K, Fuller Z, Duncan A, Holtrop G & Louis P (2009) Effect of inulin on the human gut microbiota: stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Brit J Nutr 101: 533-542.
Rose MR. Can human aging be postponed? Sci Am. 1999 Dec;281(6):106-11.
Rozan P, Nejdi A, Hidalgo S, Bisson JF, Desor D, Messaoudi M. Effects of lifelong intervention with an oligofructose-enriched inulin in rats on general health and lifespan. Br J Nutr. 2008 Dec;100(6):1192-9.
Scott KP, Martin JC, Duncan SH, Flint HJ. Prebiotic stimulation of human colonic butyrate-producing bacteria and bifidobacteria, in vitro. FEMS Microbiol Ecol. 2014 Jan;87(1):30-40.
Tap J, Mondot S, Levenez F et al. (2009) Towards the human intestinal microbiota phylogenetic core. Environ Microbiol 11: 2574-2584.
Tissenbaum, H. A. and Guarente, L. (2001). Increased dosage of a sir2 gene extends lifespan in Caenorhabditis elegans. Nature 410, 227-230
Walker AW, Ince J, Duncan SH et al. (2011) Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J 5: 220-230.
Wolever TM, Chiasson JL. Acarbose raises serum butyrate in human subjects with impaired glucose tolerance. Br J Nutr. 2000 Jul;84(1):57-61.
Witt O, Deubzer HE, Milde T, Oehme I. HDAC family: What are the cancer relevant targets? Cancer Lett. 2009 May 8;277(1):8-21.
Zhang Y, Xie Y, Berglund ED, Coate KC, He TT, Katafuchi T, Xiao G, Potthoff MJ, Wei W, Wan Y, Yu RT, Evans RM, Kliewer SA, Mangelsdorf DJ. The starvation hormone, fibroblast growth factor-21, extends lifespan in mice. Elife. 2012;1:e00065.
Zhao Y, Sun H, Lu J, Li X, Chen X, Tao D, Huang W, Huang B. Lifespan extension and elevated hsp gene expression in Drosophila caused by histone deacetylase inhibitors. J Exp Biol. 2005 Feb;208(Pt 4):697-705.
