How much selenium is optimal for health? To address this question, I’ll examine the association between circulating levels of selenium with all-cause mortality risk. Then, I’ll identify a dietary selenium intake that corresponds to optimal plasma selenium levels. Let’s have a look!
A variety of studies have investigated associations between plasma (or serum) selenium with risk of death from all causes:
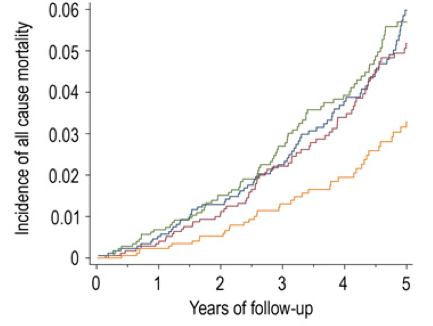
In a 9-year study of 1,389 older adults (average age, 65y) plasma selenium values less than 1.09 micromolar (uM) were associated with significantly increased all-cause mortality risk, when compared with values greater than 1.22 uM (Akbaraly et al 2005):

In a 6-year study of 1,042 older adults (average age, 76y), plasma selenium values less than 0.83 um were associated with significantly increased all-cause mortality risk, when compared with values greater than 1.0 uM (Lauretani et al. 2008):

In a 5-year study of 632 older women (average age, 74y), serum selenium values less than 1.38 uM were associated with significantly increased all-cause mortality risk, when compared with the other 3 quartiles (1.38 to 1.52, 1.53 to 1.67, and >1.68 uM; Ray et al. 2006):

In a 12-year study of 13,887 middle aged adults (average age, 45y), serum selenium values between 130-150 ng/mL (equivalent to 1.65-1.9 uM) were associated with reduced all-cause mortality risk (Bleys et al. 2006). Increased mortality risk was associated with serum selenium values less than 1.3 uM (~102 ng/mL):

These data were confirmed in the same cohort (16,008 adults) that were followed for an additional 2 years (14 years total; Goyal et al. 2013). Baseline serum selenium values greater than 1.4 uM were associated with significantly reduced all-cause mortality risk, compared with values less than1.38 uM.
Finally, in a 13-year study of 1,054 older adults (average age ~76y), elevated plasma levels of selenium (risk ratios were reported without the corresponding selenium concentration) were associated with significantly decreased all-cause mortality risk (Bates et al. 2011).
Studies that show weaker or no association between circulating values of selenium with all-cause mortality risk include Gonzalez et al. (2007) and Wei et al. (2004). In Gonzalez et al. (2007), serum selenium values greater than 1.26 uM were associated with decreased all-cause mortality risk for older women (average age, 76y), but not men, during a 4-year follow-up. However, Gonzalez et al. (2007) may have been underpowered to detect significant associations with mortality risk because of the small study size (215 total subjects). In Wei et al. (2004), a younger cohort (average age, 56y) of 1,115 subjects were followed for 15 years, and no association between serum selenium with all-cause mortality was found. However, only 4% of the population (~46 subjects) had serum selenium values greater than 1.19 uM, a finding that suggests that this study was additionally under-sized to detect significant associations.
Collectively, these studies suggest that circulating selenium values greater than at least 1.0 uM (and up to ~1.9 uM) are optimal for reducing all-cause mortality risk. What dietary intake of selenium can achieve these circulating values?
Shown below is the correlation between dietary selenium with serum selenium in 205 older adults (average age ~75y; González et al. 2006). Let’s start with the RDA selenium value for adults older than 19 years, 55 micrograms (mcg; Institute of Medicine, 2000). 55 mcg of dietary selenium is correlated with a serum selenium value of ~80 ug/L (~1 uM). In support of this correlation, a dietary selenium intake of ~47 ug/day has been shown to correlate with a circulating selenium concentration of 0.95 uM (Navarro et al. 1995). Based on the evidence already presented, eating only the RDA for selenium and achieving circulating selenium values less 1 uM would be associated with increased risk of death from all causes in 6 of the 7 studies! Based on its association with all-cause mortality risk, the RDA selenium value of 55 mcg/day is too low.

Determining which dietary selenium intake is optimal for maximally reduced all-cause mortality risk depends on how you interpret the literature. Four of previously mentioned studies showed circulating selenium values greater than 1.2 uM (95 ug/L) to be associated with reduced all-cause mortality risk. Based on the plot of González et al. (2006), ~130 ug of dietary selenium is necessary to achieve a circulating selenium concentration of 1.2 uM. Three studies showed decreased all-cause mortality risk at circulating selenium levels > 1.38 uM (110 ug/L). The dietary selenium intake that corresponds to that concentration is ~180 ug of selenium/day. Accordingly, a dietary selenium intake between 130-180 ug/day may be optimal for reducing all-cause mortality risk.
Which foods are selenium rich? Brazil nuts are the best dietary source of selenium, as 1 Brazil nut (4 g) contains 77 mcg of selenium (http://ndb.nal.usda.gov/ndb/foods/show/3641?fg=&man=&lfacet=&count=&max=&qlookup=&offset=&sort=&format=Full&reportfmt=other&rptfrm=&ndbno=&nutrient1=&nutrient2=&nutrient3=&subset=&totCount=&measureby=&_action_show=Apply+Changes&Qv=.04&Q6825=1&Q6826=1&Q6827=1). To achieve a dietary selenium intake between 130-180 mcg/day, every day I eat 1 or 2 Brazil nuts. It’s important to note that selenium toxicity can occur at intakes ~400 mcg (Food and Nutrition Board 2000), so keeping an eye on Brazil nut intake is probably a good idea.
If you’re interested, please have a look at my book!
References:
Akbaraly NT, Arnaud J, Hininger-Favier I, Gourlet V, Roussel AM, Berr C. Selenium and mortality in the elderly: results from the EVA study. Clin Chem. 2005 Nov;51(11):2117-23.
Bates CJ, Hamer M, Mishra GD. Redox-modulatory vitamins and minerals that prospectively predict mortality in older British people: the National Diet and Nutrition Survey of people aged 65 years and over. Br J Nutr. 2011 Jan;105(1):123-32.
Bleys J, Navas-Acien A, Guallar E.Serum selenium levels and all-cause, cancer, and cardiovascular mortality among US adults. Arch Intern Med. 2008 Feb 25;168(4):404-10.
Broome CS, McArdle F, Kyle JA, Andrews F, Lowe NM, Hart CA, Arthur JR, Jackson MJ. An increase in selenium intake improves immune function and poliovirus handling inadults with marginal selenium status. Am J Clin Nutr. 2004 Jul;80(1):154-62.
Food and Nutrition Board, Institute of Medicine. Selenium. Dietary reference intakes for vitamin C, vitamin E, selenium, and carotenoids. Washington, D.C.: National Academy Press; 2000:284-324.
González S, Huerta JM, Fernández S, Patterson EM, Lasheras C. Food intake and serum selenium concentration in elderly people.Ann Nutr Metab. 2006;50(2):126-31.
González S, Huerta JM, Fernández S, Patterson AM, Lasheras C. Homocysteine increases the risk of mortality in elderly individuals. Br J Nutr. 2007; 97:1138–1143.
Goyal A, Terry MB, Siegel AB. Serum antioxidant nutrients, vitamin A, and mortality in U.S. adults. Cancer Epidemiol Biomarkers Prev. 2013 Dec;22(12):2202-11.
Hurst R, Armah CN, Dainty JR, Hart DJ, Teucher B, Goldson AJ, Broadley MR, Motley AK, Fairweather-Tait SJ. Establishing optimal selenium status: results of a randomized, double-blind, placebo-controlled trial. Am J Clin Nutr. 2010 Apr;91(4):923-31.
Institute of Medicine. Food and Nutrition Board. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids. Washington, DC: National Academy Press, 2000.
Lauretani F, Semba RD, Bandinelli S, Ray AL, Ruggiero C, Cherubini A, Guralnik JM, Ferrucci L. Low plasma selenium concentrations and mortality among older community-dwelling adults: the InCHIANTI Study. Aging Clin Exp Res. 2008 Apr;20(2):153-8.
Navarro M, López H, Ruiz ML, González S, Pérez V, López MC. Determination of selenium in serum by hydride generation atomic absorption spectrometry for calculation of daily dietary intake. Sci Total Environ. 1995 Dec 15;175(3):245-52.
Ray AL, Semba RD, Walston J, Ferrucci L, Cappola AR, Ricks MO, Xue QL, Fried LP. Low serum selenium and total carotenoids predict mortality among older women living in the community: the women’s health and aging studies. J Nutr. 2006 Jan;136(1):172-6.
Swanson CA, Longnecker MP, Veillon C, Howe M, Levander OA, Taylor PR, McAdam PA, Brown CC, Stampfer MJ, Willett WC. Selenium intake, age, gender, and smoking in relation to indices of selenium status of adults residing in a seleniferous area. Am J Clin Nutr. 1990 Nov;52(5):858-62.
Wei WQ, Abnet CC, Qiao YL, Dawsey SM, Dong ZW, Sun XD, Fan JH, Gunter EW, Taylor PR, Mark SD. Prospective study of serum selenium concentrations and esophageal and gastric cardia cancer, heart disease, stroke, and total death. Am J Clin Nutr. 2004 Jan;79(1):80-5.