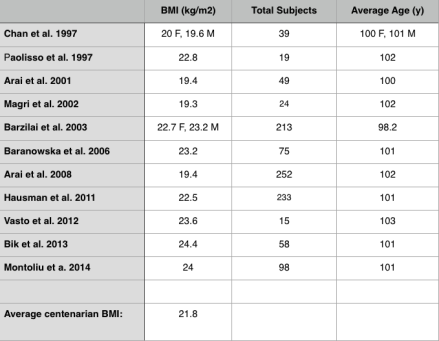
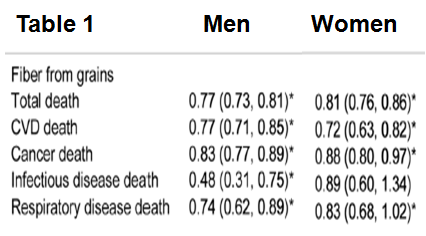
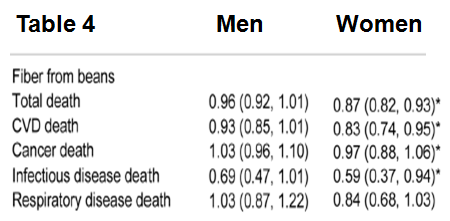
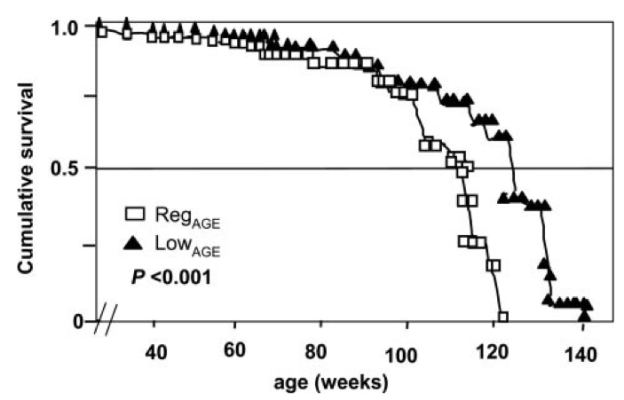
In an earlier article I wrote about how cooking foods at a high temperature (greater than 250ºF, including frying, baking, roasting and grilling) produces the neurotoxic and carcinogenic compound, acrylamide (http://voices.yahoo.com/acrylamide-chocolate-another-10217911.html?cat=5). However, the adverse effects acrylamide that I discussed were solely based on rodent studies. In this follow-up article, I’ll comprehensively discuss the evidence relating dietary acrylamide with human cancer.
Before introducing the data, it’s important to note that dietary acrylamide intake in all of the studies discussed below were calculated based on food frequency questionnaires. The highest acrylamide consuming group was approximately 40 µg/day, in comparison with low consumers of dietary acrylamide, ~10 µg /day. Without a doubt these values for dietary acrylamide intake are underestimated-for example, 1 ounce of Pringles potato chips contains 70 µg of acrylamide, and the commonly thought of as “healthier chips”, Baked Lays has 31µg/ounce (1 bag of chips).
Breast Cancer
Six large epidemiological studies (ranging from 33,000-120,000 subjects) and 1 smaller study (1500-6000 subjects) investigated the association between dietary acrylamide and breast cancer risk. Of these, 1 study, the UK Women’s Cohort Study identified a 20% significantly increased risk between acrylamide intake and premenopausal breast cancer (Burley et al. 2011). The other six studies did not show an association between acrylamide intake and breast cancer risk (Pellucchi et al. 2006, Hogervorst et al. 2007, Pedersen et al. 2009, Larsson et al. 2009, Wilson et al. 2009, Wilson et al. 2010).
Endometrial Cancer
Three large epidemiological studies have investigated the association between dietary acrylamide and endometrial cancer. In two of these studies, risk of cancer was increased by 41% and 99%, respectively (Wilson et al. 2010, Hogervorst et al. 2007). No association between dietary acrylamide intake and risk of endometrial cancer was found in the Swedish Mammography Study (Larsson et al. 2009).
Ovarian Cancer
No association between dietary acrylamide and risk of ovarian cancer was found in the small- scale Italian Cohort study, or, in 2 large-scale epidemiological studies (Pellucchi et al. 2006, Larsson et al. 2009, Wilson et al. 2010). However, a 122% increased risk for ovarian cancer in non-smokers was found in the Netherlands Cohort Study on Diet and Cancer (Hogervorst et al. 2007).
Prostate, Pancreatic, Brain Cancer
Five separate studies found no association between dietary acrylamide and risk of prostate cancer (Pellucchi et al. 2006, Hogervorst et al. 2008, Wilson et al. 2009, Larsson et al. 2009, Wilson et al. 2012). Similarly, pancreatic cancer risk is not increased (Pelucchi et al. 2011, Hogervorst et al. 2008), nor is brain cancer (Hogervorst et al. 2009), or, thyroid cancer (Schouten et al. 2009).
Esophageal cancer
One small study (987 subjects) found a 23% increased risk for esophageal cancer, and an 88% increased risk in those with a BMI greater than 25. In two other studies (Pellucchi et al. 2006, Hogervorst et al. 2008), no association between dietary acrylamide and esophageal cancer was found.
Head-neck cancer
Increased risk for oral-cavity cancer in female non-smokers in a large study (121,000 subjects; Schouten et al. 2009) was found. No association for oral cavity, pharynx or larynx cancer in a smaller study (1500-6000 subjects; Pellucchi et al. 2006)
Kidney Cancer
Although risk of kidney cancer was significantly increased by 59%, it appears as if this data was skewed by smokers. In non-smokers, risk of kidney cancer was not significant (Pellucchi et al. 2006). No association between dietary acrylamide and risk of kidney cancer was also identified in three additional studies (Mucci et al. 2003, Mucci et al. 2004, Pellucchi et al. 2007).
Gastric, Colon, Rectal cancer
A small study with 1129 subjects found a 40% decreased risk of large bowel cancer (Mucci et al. 2003). Four studies have not found a similar association (Pellucchi et al. 2006, Mucci et al. 2006, Hogervorst et al. 2008, Larsson et al. 2009).
Lung Cancer
A 55% decreased risk of lung cancer, in women was identified by Hogervorst et al. (2009).
Bladder cancer
Significant only in smokers, as 15+ cigarettes/day significantly increased risk of bladder cancer in those with the highest dietary acrylamide intake, relative to the lowest intake (Hogervorst et al. 2008).
Blood cancer
Multiple myeloma and follicular myeloma were found to be significantly increased by 14% and 28% for every 10 µg increment in dietary acrylamide (Bongers et al. 2012).
Conclusions
The easy interpretation of scientific studies is that if six studies show no effect and one study shows a positive effect, that the no effect-data is the real answer. For example, in the case of breast cancer, six studies showed no effect, whereas one study showed a significant association between acrylamide and premenopausal breast cancer. Should we conclude that there is no risk for breast cancer? As I mentioned earlier, it is likely that total dietary acrylamide intake was underestimated, and therefore, it is my opinion that none of the 25 studies should have shown an association between acrylamide and cancer. Therefore, that there was indeed a significant association for breast cancer with potentially underestimated acrylamide values is significant. Also, dietary acrylamide was shown to be significantly associated with myeloma, head-neck cancer, esophageal cancer, endometrial cancer and ovarian cancer. Paradoxically, dietary acrylamide reduced risk of lung and large bowel cancer.
What should someone who is interested in optimal health do with this information? Knowing that dietary acrylamide is indeed significantly associated with increased risk of human cancers, I would reduce or eliminate cooking food at a high temperature. I have!
If you’re interested, please have a look at my book!
References:
Bongers ML, Hogervorst JG, Schouten LJ, Goldbohm RA, Schouten HC, van den Brandt PA. Dietary acrylamide intake and the risk of lymphatic malignancies: the Netherlands Cohort Study on diet and cancer. PLoS One. 2012;7(6):e38016.
Burley VJ, Greenwood DC, Hepworth SJ, Fraser LK, de Kok TM, van Breda SG, Kyrtopoulos SA, Botsivali M, Kleinjans J, McKinney PA, Cade JE. Dietary acrylamide intake and risk of breast cancer in the UK women’s cohort. Br J Cancer. 2010 Nov 23;103(11):1749-54.
Hogervorst JG, Schouten LJ, Konings EJ, Goldbohm RA, van den Brandt PA. A prospective study of dietary acrylamide intake and the risk of endometrial, ovarian, and breast cancer. Cancer Epidemiol Biomarkers Prev. 2007 Nov;16(11):2304-13.
Hogervorst JG, Schouten LJ, Konings EJ, Goldbohm RA, van den Brandt PA. Dietary acrylamide intake and the risk of renal cell, bladder, and prostate cancer. Am J Clin Nutr. 2008 May;87(5):1428-38
Hogervorst JG, Schouten LJ, Konings EJ, Goldbohm RA, van den Brandt PA. Dietary acrylamide intake is not associated with gastrointestinal cancer risk. J Nutr. 2008 Nov;138(11):2229-36.
Hogervorst JG, Schouten LJ, Konings EJ, Goldbohm RA, van den Brandt PA. Lung cancer risk in relation to dietary acrylamide intake. J Natl Cancer Inst. 2009 May 6;101(9):651-62.
Hogervorst JG, Schouten LJ, Konings EJ, Goldbohm RA, van den Brandt PA. Dietary acrylamide intake and brain cancer risk. Cancer Epidemiol Biomarkers Prev. 2009 May;18(5):1663-6.
Larsson SC, Akesson A, Wolk A. Long-term dietary acrylamide intake and breast cancer risk in a prospective cohort of Swedish women. Am J Epidemiol. 2009 Feb 1;169(3):376-81.
Larsson SC, Håkansson N, Akesson A, Wolk A. Long-term dietary acrylamide intake and risk of endometrial cancer in a prospective cohort of Swedish women. Int J Cancer. 2009 Mar 1;124(5):1196-9.
Larsson SC, Akesson A, Bergkvist L, Wolk A. Dietary acrylamide intake and risk of colorectal cancer in a prospective cohort of men. Eur J Cancer. 2009 Mar;45(4):513-6.
Larsson SC, Akesson A, Wolk A. Long-term dietary acrylamide intake and risk of epithelial ovarian cancer in a prospective cohort of Swedish women. Cancer Epidemiol Biomarkers Prev. 2009 Mar;18(3):994-7.
Larsson SC, Akesson A, Wolk A. Dietary acrylamide intake and prostate cancer risk in a prospective cohort of Swedish men. Cancer Epidemiol Biomarkers Prev. 2009 Jun;18(6):1939-41.
Lin Y, Lagergren J, Lu Y. Dietary acrylamide intake and risk of esophageal cancer in a population-based case-control study in Sweden. Int J Cancer. 2011 Feb 1;128(3):676-81.
Mucci LA, Dickman PW, Steineck G, Adami HO, Augustsson K. Dietary acrylamide and cancer of the large bowel, kidney, and bladder: absence of an association in a population-based study in Sweden. Br J Cancer. 2003 Jan 13;88(1):84-9.
Mucci LA, Lindblad P, Steineck G, Adami HO. Dietary acrylamide and risk of renal cell cancer. Int J Cancer. 2004 May 1;109(5):774-6.
Mucci LA, Adami HO, Wolk A. Prospective study of dietary acrylamide and risk of colorectal cancer among women. Int J Cancer. 2006 Jan 1;118(1):169-73.
Pedersen GS, Hogervorst JG, Schouten LJ, Konings EJ, Goldbohm RA, van den Brandt PA. Dietary acrylamide intake and estrogen and progesterone receptor-defined postmenopausal breast cancer risk. Breast Cancer Res Treat. 2010 Jul;122(1):199-210.
Pelucchi C, Galeone C, Levi F, Negri E, Franceschi S, Talamini R, Bosetti C, Giacosa A, La Vecchia C. Dietary acrylamide and human cancer. Int J Cancer. 2006 Jan 15;118(2):467-71.
Pelucchi C, Galeone C, Dal Maso L, Talamini R, Montella M, Ramazzotti V, Negri E, Franceschi S, La Vecchia C. Dietary acrylamide and renal cell cancer. Int J Cancer. 2007 Mar 15;120(6):1376-7.
Pelucchi C, Galeone C, Talamini R, Negri E, Polesel J, Serraino D, La Vecchia C. Dietary acrylamide and pancreatic cancer risk in an Italian case–control study. Ann Oncol. 2011 Aug;22(8):1910-5.
Schouten LJ, Hogervorst JG, Konings EJ, Goldbohm RA, van den Brandt PA. Dietary acrylamide intake and the risk of head-neck and thyroid cancers: results from the Netherlands Cohort Study. Am J Epidemiol. 2009 Oct 1;170(7):873-84.
Wilson KM, Mucci LA, Cho E, Hunter DJ, Chen WY, Willett WC. Dietary acrylamide intake and risk of premenopausal breast cancer. Am J Epidemiol. 2009 Apr 15;169(8):954-61.
Wilson KM, Bälter K, Adami HO, Grönberg H, Vikström AC, Paulsson B, Törnqvist M, Mucci LA. Acrylamide exposure measured by food frequency questionnaire and hemoglobin adduct levels and prostate cancer risk in the Cancer of the Prostate in Sweden Study. Int J Cancer. 2009 May 15;124(10):2384-90.
Wilson KM, Mucci LA, Rosner BA, Willett WC. A prospective study on dietary acrylamide intake and the risk for breast, endometrial, and ovarian cancers. Cancer Epidemiol Biomarkers Prev. 2010 Oct;19(10):2503-15.
Wilson KM, Giovannucci E, Stampfer MJ, Mucci LA. Dietary acrylamide and risk of prostate cancer. Int J Cancer. 2012 Jul 15;131(2):479-87.